Subterranean Biology Subterranean Biology 11: 45–53, doi: 10.3897/subtbiol.11.5966New
Zospeum species (Gastropoda, Ellobioidea, Carychiidae) from 980 m depth in the Lukina Jama – Trojama cave system (Velebit Mts., Croatia)Alexander M. Weigand 1,2,† 1 Department of Phylogeny and Systematics, Institute for Ecology, Evolution and Diversity, Biosciences, Goethe-University Frankfurt, Max-von-Laue Straße 13, 60438 Frankfurt am Main, Germany 2 Croatian Biospeleological Society, Demetrova 1, 10000 Zagreb, Croatia †
http://zoobank.org/C15902F9-515F-4D53-94A7-59058492A569 Corresponding author: Alexander M. Weigand (
email address)
Academic editor: Oana Moldovanreceived 12 June 2013 | accepted 14 August 2013 | Published 30 August 2013
(C) 2013 Alexander M. Weigand. This is an open access article distributed under the terms of the
Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Weigand AM (2013) New
Zospeum species (Gastropoda, Ellobioidea, Carychiidae) from 980 m depth in the Lukina Jama–Trojama cave system (Velebit Mts., Croatia). Subterranean Biology 11: 45–53. doi:
10.3897/subtbiol.11.5966 AbstractA new species of the eutroglobiont gastropod taxon
Zospeum Bourguignat, 1856 is described.
Zospeum tholussum sp. n. is characterized based on a population from the Lukina Jama–Trojama cave system (Velebit Mts., Croatia). A single living specimen occurred at 980 m depth. The species is morphologically related to
Zospeum amoenum (Frauenfeld, 1856), but can be readily distinguished from the latter by the presence of a weak columellar fold and its dome-like structured 2
nd whorl. DNA barcoding is capable to clearly delineate
Zospeum tholussum from other
Zospeum spp. as well.KeywordsDNA barcoding, cryptic species, biospeleology, eutroglobiont gastropod, cave-dwelling species, microgastropoda IntroductionTaxonomic research distinguishes eleven morphospecies (no subspecies considered) of the microgastropod taxon
Zospeum Bourguignat, 1856 (
Ellobioidea,
Carychiidae) for the region of the Dinaric Alps (Bole 1974, Slapnik and Ozimec 2004, de Jong 2012). Their taxonomic history has been shaped by confusing and sometimes contradicting (sub-)species assignments, primarily as a result of the general problem to distinguish intraspecific and interspecific shell variability (Bole 1974), which for decades was the main source of taxonomic information. Recently, Weigand et al. (2011) implemented DNA barcodes to molecularly identify species (i.e. DNA barcoding) based on a fragment of the mitochondrial-encoded (mtDNA) Cytochrome C Oxidase Subunit I (COI) locus (Hajibabaei et al. 2007). In a subsequent study and by addressing the evolutionary history of
Carychiidae, Weigand et al. (2013) revealed several morphologically unrecognized (cryptic)
Zospeum species and demonstrated the suitability of molecular data for taxonomic purposes in this group.
All known
Zospeum have lost visual orientation and are considered true eutroglobionts (see Sket 2008). Information on their biology is scarce, but species seem to possess a limited active movement (Slapnik 2001) and to prefer muddy and permanently wet subterranean microhabitats (Jochum et al. 2012), as generally found along the drainage system of a cave. Weigand et al. (2013) proposed that dispersal may primarily occur via passive transportation (e.g. by water or larger mammals).
Here, a new
Zospeum population from the Lukina Jama–Trojama cave system situated in the Velebit mountain range of Croatia is characterized (Fig. 1). On the basis of molecular and conchological data, a new species from 980 m depth–
Zospeum tholussum Weigand, sp. n.–is described.
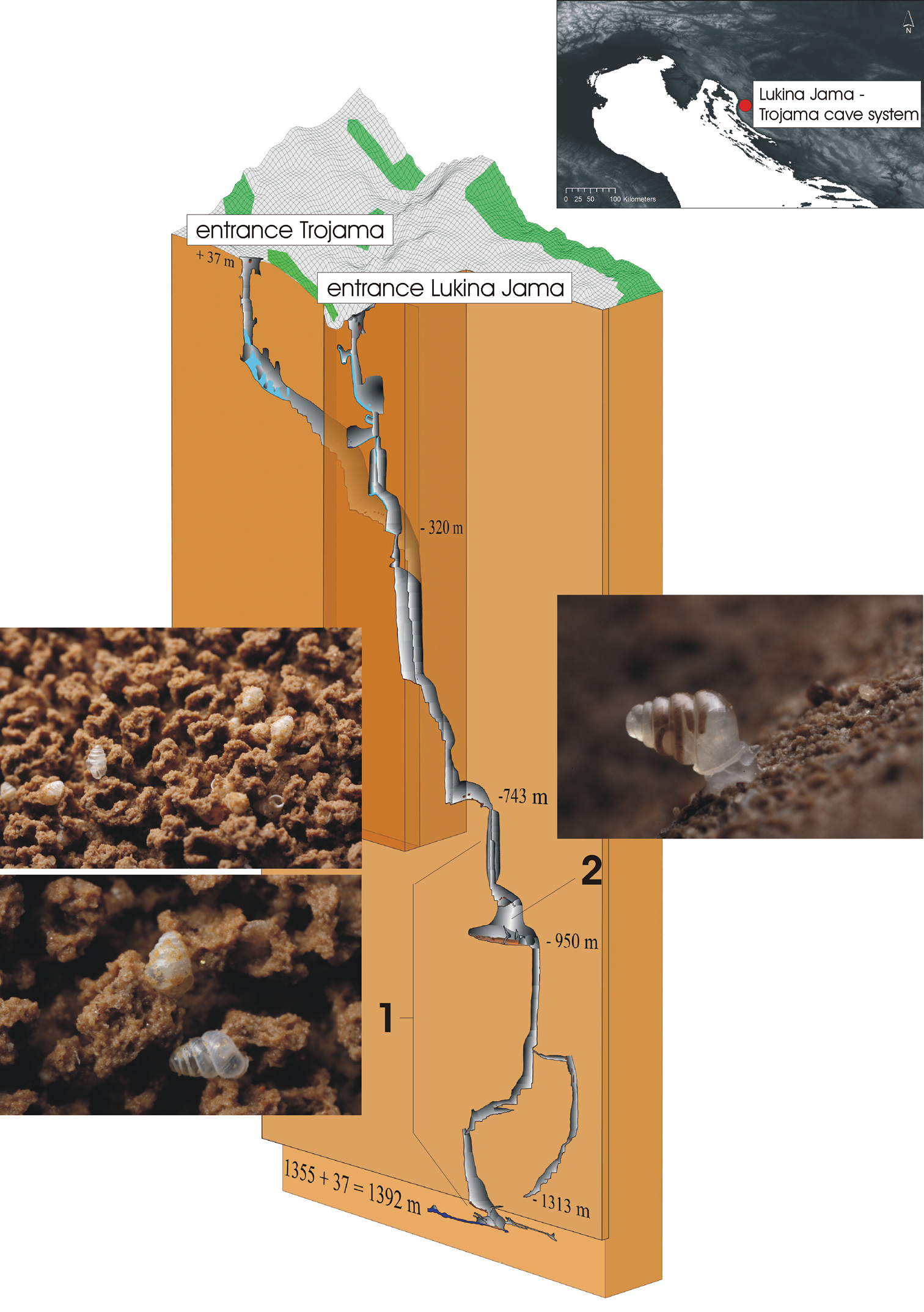
Figure 1. The Lukina Jama–Trojama cave system. Overview of the geographical position and 3D cave cross-section. In the latter, the region of collected shells (1) and the collection site of the living specimen of
Zospeum tholussum (2) are indicated. The 3D cross-section was provided by D. Bakšić et al. (2010), Croatian Speleological Server,
http://www.speleologija.hr/lukinajama. Photos were taken by J. Bedek.
Figure 1. The Lukina Jama–Trojama cave system. Overview of the geographical position and 3D cave cross-section. In the latter, the region of collected shells (1) and the collection site of the living specimen of
Zospeum tholussum (2) are indicated. The 3D cross-section was provided by D. Bakšić et al. (2010), Croatian Speleological Server,
http://www.speleologija.hr/lukinajama. Photos were taken by J. Bedek. Methods Conchological measurementsSpecimen shells were measured by taking individual images with 456 pixels corresponding to 1 mm. The total shell height (SH), shell width (SW), aperture height (AH), aperture width (AW) and the number of whorls were measured. Moreover, ratios for SH/SW and AH/AW were calculated. Non-invasive DNA isolation, PCR and sequencingSpecimen shells of live-collected
Zospeum tholussum sp. n. (single specimen) and
Zospeum amoenum (Frauenfeld, 1856) (two specimens) were kept intact by using a protocol for a non-invasive DNA isolation method originally described in Schizas et al. (1997) and partially modified after Böttger-Schnack and Machida (2011). Each specimen was cleaned and then dried in a 0.2 mL PCR-tube by raising the temperature once up to 94 °C. A volume of 9 µL ddH
2O and 1 µL 10x PCR-buffer were added. The mixture was heated for 2 min at 94 °C; 1.3 µL Proteinase K were added and the solution homogenized by gentle mixing. An incubation step was performed for 15 min at 55 °C followed by 10 min at 70 °C. The incubation was repeated once. 10 µL of Gene Releaser (Bioventures Inc.) were added and the following thermocycler protocol used: 30 s at 65 °C, 30 s at 8 °C, 1.5 min at 65 °C, 3 min at 97 °C, 1 min at 8 °C, 3 min at 65 °C, 1 min at 97 °C, 1 min at 65 °C, 5 min at 80 °C and storage at 4 °C. The reaction mixture including the intact shell was centrifuged for 1 min using a table centrifuge. The clean phase containing the DNA was collected and transferred to another 0.2 mL PCR-tube. The shell was cleaned from the remains of the Gene Releaser chemicals by alternate rinsing in water and ethanol. 10–15 µL of AE Buffer (DNeasy Kit, Qiagen) were added to the isolated DNA.
PCR conditions were the same as described in Weigand et al. (2011), sequencing was performed at Source BioScience (LifeSciences, Berlin, Germany) following the companies instructions. DNA barcodingDNA barcodes, chromatograms, images, geographic data and further information of the genetically analysed
Zospeum tholussum (BOLD-ID
BARCA210-13) and
Zospeum amoenum (BOLD-IDs
BARCA211-13 and
BARCA212-13) specimens are deposited in the Barcode of Life Database (BOLD) (Ratnasingham and Hebert 2007) in the project “Barcoding Carychiid Microsnails” [BARCA]. The obtained COI-sequences were compared with other
Zospeum species in BOLD. Results Family
Carychiidae Jeffreys, 1830 Genus
Zospeum Bourguignat, 1856
Zospeum tholussum Weigand, sp. n.
http://zoobank.org/A3B380AD-B918-451B-866D-7FA59642A733http://species-id.net/wiki/Zospeum_tholussumFigs 1–4 Material examined.A single living specimen was collected on 31.07.2010 (leg. Jana Bedek), which is designated as the holotype specimen (BOLD-ID
BARCA210-13) (Fig. 1, life image; Fig. 2, upper row). Eight shells comprise paratype specimens, of which one was partially broken and used to investigate the form of the columellar fold. Shells were collected on several days during the caving expedition from 29.07. - 03.08.2010 (Fig. 2). All investigated specimens are deposited in the Forschungsinstitut und Naturmuseum Senckenberg, Frankfurt am Main, Germany (museum voucher SMF 341633).

Figure 2. Holotype and paratypes of
Zospeum tholussum. The holotype (former living specimen) is marked with a solid line; five paratype specimens (shells) are surrounded by dotted lines.
Figure 2. Holotype and paratypes of
Zospeum tholussum. The holotype (former living specimen) is marked with a solid line; five paratype specimens (shells) are surrounded by dotted lines.Diagnosis.The general shape of
Zospeum tholussum sp. n. resembles larger specimens of
Zospeum amoenum (Frauenfeld, 1856) from which it can be best delineated by the dome-like structured 2
nd whorl (Fig. 2), the weak but present columellar fold (Fig. 3) and by means of their DNA barcodes (11.7 %–12.1 % genetic p-distance between both species).Description.The eight conchologically investigated specimens (the holotype and seven paratypes) demonstrate moderate shell variability (Table 1). Shells are generally smooth, very thin and fragile. Fresh shells or those of living specimens are translucent, older ones adopt a milky white color. The number of whorls are 5–6, SH ranges from 1.41–1.81 mm, SW from 0.93–1.12 mm, AH from 0.44–0.54 mm and AW from 0.38–0.46 mm. The ratio of SH/SW is between 1.34–1.62 and AH/AW between 1.05–1.30. The 2
nd whorl is remarkably enlarged possessing a height of 2/3–5/6 the height of the 3
rd + 4
th whorl. The columellar is only weakly developed (Fig. 3) and no teeth (i.e. parietalis, palatalis or columellaris) are present. The DNA barcode of the holotype, when compared with other DNA barcodes of
Zospeum species deposited in BOLD, shows its lowest interspecific genetic distance to
Zospeum pretneri Bole, 1960 (5.6 %; BOLD-ID:
BARCA120-10). This result is well above the barcoding gap of 3.2 % for
Carychiidae, which is suitable to separate between-species (> 3.2 %, interspecific) and within-species genetic diversity (< 3.2 %, intraspecific) in
Zospeum (Weigand et al. 2011, 2013).Table 1. Conchological measurements for
Zospeum tholussum. Seven paratypes (Ind. 1–7) and the holotype specimen (Ind.

were measured. All measurements are given in millimeter. Additionally, ratios of shell height / shell width (sh/sw) and aperture height / aperture width (ah/aw) were calculated. The range of variability is given for all parameters.
Ind. sh/sw ah/aw shell height shell width aperture height aperture width |
| 1 | 1.60 | 1.16 | 1.49 | 0.93 | 0.44 | 0.38 |
| 2 | 1.42 | 1.30 | 1.54 | 1.08 | 0.52 | 0.40 |
| 3 | 1.46 | 1.16 | 1.41 | 0.97 | 0.46 | 0.39 |
| 4 | 1.34 | 1.05 | 1.44 | 1.07 | 0.47 | 0.44 |
| 5 | 1.50 | 1.31 | 1.63 | 1.09 | 0.54 | 0.41 |
| 6 | 1.53 | 1.23 | 1.61 | 1.05 | 0.51 | 0.41 |
| 7 | 1.62 | 1.15 | 1.81 | 1.12 | 0.53 | 0.46 |
| 8 (holotype) | 1.48 | 1.12 | 1.47 | 0.99 | 0.44 | 0.39 |
| range | 1.34–1.62 | 1.05–1.30 | 1.41–1.81 | 0.93–1.12 | 0.44–0.54 | 0.38–0.46 |
Ind. sh/sw ah/aw shell height shell width aperture height aperture width |
| 1 | 1.60 | 1.16 | 1.49 | 0.93 | 0.44 | 0.38 |
| 2 | 1.42 | 1.30 | 1.54 | 1.08 | 0.52 | 0.40 |
| 3 | 1.46 | 1.16 | 1.41 | 0.97 | 0.46 | 0.39 |
| 4 | 1.34 | 1.05 | 1.44 | 1.07 | 0.47 | 0.44 |
| 5 | 1.50 | 1.31 | 1.63 | 1.09 | 0.54 | 0.41 |
| 6 | 1.53 | 1.23 | 1.61 | 1.05 | 0.51 | 0.41 |
| 7 | 1.62 | 1.15 | 1.81 | 1.12 | 0.53 | 0.46 |
| 8 (holotype) | 1.48 | 1.12 | 1.47 | 0.99 | 0.44 | 0.39 |
| range | 1.34–1.62 | 1.05–1.30 | 1.41–1.81 | 0.93–1.12 | 0.44–0.54 | 0.38–0.46 |

Figure 3. Columellar fold views of
Zospeum tholussum. The columellar fold of the broken paratype specimen shell is shown in clockwise rotation.
Figure 3. Columellar fold views of
Zospeum tholussum. The columellar fold of the broken paratype specimen shell is shown in clockwise rotation.Habitat.The single living specimen was found in an unnamed large chamber at 980 m depth (85 m long, 70 m wide) with lots of stones, rocks and sand (Fig. 1, right site). A temporal small stream of running water was present close to the collecting site. Air temperature was between 3.3–3.5 °C (depending on the measurement device), water temperature 5.1 °C and air humidity 100 %. Shells were observed beginning from 800 m depth till the bottom of the cave. Shells were generally found on layers of mud (Fig. 1, left site). The first 200 m of the entrance passage of Lukina Jama are permanently covered by varying levels of snow and ice.Etymology.The Latin word
tholus means dome or cupola and refers to the remarkable dome-like shape of the 2
nd whorl.Remarks.In addition to the newly described
Zospeum tholussum, a second
Zospeum species is present in the Lukina Jama–Trojama cave system (Fig. 4C). This species can be differentiated from
Zospeum tholussum by the presence of a tooth, its general shell shape, more prominent columellar fold and absence of the characteristic dome-like structured 2
nd whorl. Because only shells were found, no DNA barcodes are available for this species.

Figure 4.
Zospeum species.
A Zospeum tholussum holotype
B Zospeum pretneri from
locus typicus, which demonstrates the lowest genetic distance to
Zospeum tholussum C Second
Zospeum sp. from the Lukina Jama–Trojama cave system.
Figure 4.
Zospeum species.
A Zospeum tholussum holotype
B Zospeum pretneri from
locus typicus, which demonstrates the lowest genetic distance to
Zospeum tholussum C Second
Zospeum sp. from the Lukina Jama–Trojama cave system.So far,
Zospeum tholussum is only known from the Lukina Jama–Trojama cave system. However, this cave system is situated in the distribution range of the morphologically related species
Zospeum amoenum (Frauenfeld, 1856), which inhabits caves of the West Balkan of North Slovenia, West Croatia, Bosnia and Herzegovina and South to Montenegro (Bole 1974). Absolon (1916) named but did not describe another morphologically related species -
Zospeum troglobalcanicum. Since Weigand et al. (2013) revealed that
Zospeum spp. with an inferred large distribution range are particularly prone to host additional, morphologically unrecognized species, some of the distribution records for
Zospeum amoenum may very well refer to
Zospeum tholussum. Moreover, the intraspecific genetic and conchological variability of the new species should be investigated in greater detail, thus to judge about the taxonomic information content of the diagnostic characters applied within this description.
The preliminary information on the habitat of
Zospeum tholussum is in congruence with previous findings for
Zospeum. Interestingly, the grazing-labyrinth-like structure, in which most of the shells were embedded (Fig. 1, left site), has been already observed during caving expeditions in Northern Spain (Jochum et al. 2012). Finally, the presence of a temporal stream at the collecting site of the living specimen further supports a passive dispersal scenario as suggested by Weigand et al. (2013). AcknowledgementsI thank Jana Bedek and all members of the HBSD (Zagreb, Croatia) for their permanent biospeleological effort and the samples, Adrienne Jochum (Frankfurt, Germany) and Rajko Slapnik (Ljubljana, Slovenia) for their taxonomic assistance and Yasunori Kano (Tokyo, Japan) for his introduction to the method of non-invasive DNA isolation. The research on the adjustment of this method to carychiid microgastropods carried out at the AORI (University of Tokyo, Japan) was supported by the Japan Society for the Promotion of Science (JSPS). The 3D cross-section of the Lukina Jama–Trojama cave system was kindly provided by D. Bakšić et al. (2010), Croatian Speleological Server,
http://www.speleologija.hr/lukinajama. The collecting permit for the Croatian Biospeleological Society (HBSD) was issued by the Croatian Ministry of Culture, dated 24 June 2010, no. 538-08-01-01/3-10-02. ReferencesAbsolon K (1916) Z výzkumnýh cest po kraseh balkánu. Zlatá Praha 33.Absolon K (1916) Z výzkumnýh cest po kraseh balkánu. Zlatá Praha 33.Bole J (1960) Novi vrsti iz rodu
Zospeum Bourg. (Gastropoda). Biološki vestnik 7(1): 61-64.Bole J (1960) Novi vrsti iz rodu
Zospeum Bourg. (Gastropoda). Biološki vestnik 7(1): 61-64.Bole J (1974) Rod
Zospeum Bourguignat 1856 (Gastropoda, Ellobiidae) v Jugoslaviji. Slovenska Akademija Znanosti in Umetnosti. Razprave 17(5): 1-43.Bole J (1974) Rod
Zospeum Bourguignat 1856 (Gastropoda, Ellobiidae) v Jugoslaviji. Slovenska Akademija Znanosti in Umetnosti. Razprave 17(5): 1-43.Böttger-Schnack R, Machida RJ (2011) Comparison of morphological and molecular traits for species identification and taxonomic grouping of oncaeid copepods. Hydrobiologia 666(1): 111-125. doi:
10.1007/s10750-010-0094-1Böttger-Schnack R, Machida RJ (2011) Comparison of morphological and molecular traits for species identification and taxonomic grouping of oncaeid copepods. Hydrobiologia 666(1): 111-125. doi:
10.1007/s10750-010-0094-1Bourguignat JR (1856) Aménités malacologiques. Revue et Magasin de Zoologie pure et appliquée (2) 8: 499–516.Bourguignat JR (1856) Aménités malacologiques. Revue et Magasin de Zoologie pure et appliquée (2) 8: 499–516.de Jong YSDM (2012) Fauna Europaea version 2.5. Web Service available online at
http://www.faunaeur.orgde Jong YSDM (2012) Fauna Europaea version 2.5. Web Service available online at
http://www.faunaeur.orgFrauenfeld G (1856) Die Gattung
Carychium. Sitzungsberichte der Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse 19(1): 70-93.Frauenfeld G (1856) Die Gattung
Carychium. Sitzungsberichte der Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse 19(1): 70-93.Hajibabaei M, Singer GA, Hebert PD, Hickey DA (2007) DNA barcoding: how it complements taxonomy, molecular phylogenetics and population genetics. Trends in Genetics 23(4): 167-172. doi:
10.1016/j.tig.2007.02.001Hajibabaei M, Singer GA, Hebert PD, Hickey DA (2007) DNA barcoding: how it complements taxonomy, molecular phylogenetics and population genetics. Trends in Genetics 23(4): 167-172. doi:
10.1016/j.tig.2007.02.001Jeffreys JG (1830) A synopsis on the testaceous pneumonobranchous Mollusca of Great Britain. Transactions of the Linnean Society of London 16(2): 323-392. doi:
10.1111/j.1095-8339.1829.tb00139.xJeffreys JG (1830) A synopsis on the testaceous pneumonobranchous Mollusca of Great Britain. Transactions of the Linnean Society of London 16(2): 323-392. doi:
10.1111/j.1095-8339.1829.tb00139.xJochum A, Weigand AM, Slapnik R, Valentinčič J, Prieto CE (2012) The microscopic ellobioid,
Zospeum Bourguignat, 1856 (Pulmonata, Ellobioidea, Carychiidae) makes a big debut in Basque Country and the province of Burgos (Spain). MalaCo 8: 400-403.Jochum A, Weigand AM, Slapnik R, Valentinčič J, Prieto CE (2012) The microscopic ellobioid,
Zospeum Bourguignat, 1856 (Pulmonata, Ellobioidea, Carychiidae) makes a big debut in Basque Country and the province of Burgos (Spain). MalaCo 8: 400-403.Ratnasingham S, Hebert PD (2007) BOLD: The Barcode of Life Data System (
http://www.barcodinglife.org). Molecular Ecology Notes 7(3): 355-364. doi:
10.1111/j.1471-8286.2007.01678.xRatnasingham S, Hebert PD (2007) BOLD: The Barcode of Life Data System (
http://www.barcodinglife.org). Molecular Ecology Notes 7(3): 355-364. doi:
10.1111/j.1471-8286.2007.01678.xSchizas NV, Street GT, Coull BC, Chandler GT, Quattro JM (1997) An efficient DNA extraction method for small metazoans. Molecular Marine Biology and Biotechnology 6(4): 381.Schizas NV, Street GT, Coull BC, Chandler GT, Quattro JM (1997) An efficient DNA extraction method for small metazoans. Molecular Marine Biology and Biotechnology 6(4): 381.Sket B (2008) Can we agree on an ecological classification of subterranean animals?. Journal of Natural History 42(21–22): 1549-1563. doi:
10.1080/00222930801995762Sket B (2008) Can we agree on an ecological classification of subterranean animals?. Journal of Natural History 42(21–22): 1549-1563. doi:
10.1080/00222930801995762Slapnik R (2001) Activity and movements of
Zospeum isselianum Pollonera 1886 (Gastropoda, Pulmonata, Carychiidae) in a cave in the Kamniške-Savinjske Alps (Slovenia). Natura Croatica 10(3): 153-162.Slapnik R (2001) Activity and movements of
Zospeum isselianum Pollonera 1886 (Gastropoda, Pulmonata, Carychiidae) in a cave in the Kamniške-Savinjske Alps (Slovenia). Natura Croatica 10(3): 153-162.Slapnik R, Ozimec R (2004) Distribution of the genus
Zospeum Bourguignat 1856 (Gastropoda, Pulmonata, Ellobiidae) in Croatia. Natura Croatica 13(2): 115-135.Slapnik R, Ozimec R (2004) Distribution of the genus
Zospeum Bourguignat 1856 (Gastropoda, Pulmonata, Ellobiidae) in Croatia. Natura Croatica 13(2): 115-135.Weigand AM, Jochum A, Pfenninger M, Steinke D, Klussmann-Kolb A (2011) A new approach to an old conundrum - DNA barcoding sheds new light on phenotypic plasticity and morphological stasis in microsnails (Gastropoda, Pulmonata, Carychiidae). Molecular Ecology Resources 11(2): 255-265. doi:
10.1111/j.1755-0998.2010.02937.xWeigand AM, Jochum A, Pfenninger M, Steinke D, Klussmann-Kolb A (2011) A new approach to an old conundrum - DNA barcoding sheds new light on phenotypic plasticity and morphological stasis in microsnails (Gastropoda, Pulmonata, Carychiidae). Molecular Ecology Resources 11(2): 255-265. doi:
10.1111/j.1755-0998.2010.02937.xWeigand AM, Jochum A, Slapnik R, Schnitzler J, Zarza E, Klussmann-Kolb A (2013) Evolution of microgastropods (Ellobioidea, Carychiidae): integrating taxonomic, phylogenetic and evolutionary hypotheses. BMC Evolutionary Biology 13(1): 18. doi:
10.1186/1471-2148-13-18